
Arxius de Miscel·lània Zoològica. Volumen 18 (2020) Páginas: 89-100
Bat species richness and activity in Dghoumes National Park (Southwest Tunisia): a preliminary survey
Dalhoumi, R., Aissa, P., Beyrem, H., Aulagnier, S.
DOI: https://doi.org/10.32800/amz.2020.18.0089Descargar
PDFCita
Dalhoumi, R., Aissa, P., Beyrem, H., Aulagnier, S., 2020. Bat species richness and activity in Dghoumes National Park (Southwest Tunisia): a preliminary survey. Arxius de Miscel·lània Zoològica, 18: 89-100, DOI: https://doi.org/10.32800/amz.2020.18.0089-
Fecha de recepción:
- 29/01/2020
-
Fecha de aceptación:
- 17/07/2020
-
Fecha de publicación:
- 03/08/2020
-
Compartir
-


-
Visitas
- 3926
-
Descargas
- 885
Abstract
Bat species richness and activity in Dghoumes National Park (Southwest Tunisia): a preliminary survey
Bat fauna in eight of the main habitat types of Dghoumes National Park was inventoried using mist-netting, acoustic detection and roost search. Bats were active at night and recorded near water bodies and street lamps. We recorded the echolocation calls of six bat species: Eptesicus isabellinus, Pipistrellus kuhlii, Vansonia rueppellii, Asellia tridens, Tadarida teniotis and Rhinopoma cystops. Two bat colonies containing 111 individuals of R. cystops were found roosting in Jebel Morra cave and 54 individuals of A. tridens were found roosting in the ceiling of the Ecomuseum. Due to potential disturbance by visitors to the museum, we suggest strengthening management practices to ensure the usage of this roosting site in order to promote the conservation of A. tridens.
Data published through GBIF (Doi: 10.15470/0u03uz)
Key words: Chiroptera, Acoustic, Night activity, Water bodies, Northwest Africa
Resumen
Riqueza de especies y actividad de murciélagos en el Parque Nacional de Dghoumes (suroeste de Túnez): estudio preliminar
Se estudió la fauna de murciélagos en ocho de los principales tipos de hábitats del Parque Nacional de Dghoumes utilizando redes de niebla, detección acústica y búsqueda de refugios. Los murciélagos estuvieron activos durante la noche y se localizaron en la proximidad de masas de agua y farolas de alumbrado público. Se registraron seis especies de murciélagos: Eptesicus isabellinus, Pipistrellus kuhlii, Vansonia rueppellii, Asellia tridens, Tadarida teniotis y Rhinopoma cystops. Se localizaron dos colonias, una de R. cystops con 111 individuos colgados en la cueva de Jebel Morra y otra con 54 individuos de A. tridens en el techo del Ecomuseo. Ante la posible perturbación causada por los visitantes del museo, sugerimos reforzar las medidas de gestión del público para garantizar el uso de este lugar de descanso de los murciélagos, lo que impactaría positivamente en la conservación de A. tridens.
Datos publicados en GBIF (Doi: 10.15470/0u03uz)
Palabras clave: Chiroptera, Detección acústica, Actividad nocturna, Masas de agua, Noroeste de África
Resum
Riquesa d’espècies i activitat de ratpenats al Parc Nacional de Dghoumes (sud-oest de Tunísia): estudi preliminar
Es va portar a terme un estudi de la fauna de ratpenats en vuit dels principals tipus d’hàbitats del Parc Nacional de Dghoumes utilitzant xarxes de boira, detecció acústica i cerca de refugis. Els ratpenats van estar actius durant la nit i es van localitzar prop de masses d’aigua i fanals d’enllumenat públic. Es van registrar sis espècies de ratpenats: Eptesicus isabellinus, Pipistrellus kuhlii, Vansonia rueppellii, Asellia tridens, Tadarida teniotis i Rhinopoma cystops. Es van localitzar dues colònies, una de R. cystops amb 111 individus penjats a la cova de Jebel Morra i una altra amb 54 individuos d’A. tridens al sostre de l’Ecomuseu. Davant la possible pertorbació causada pels visitants del museu, suggerim reforçar les mesures de gestió del públic per garantir l’ús d’aquest lloc de descans dels ratpenats, la qual cosa impactaria positivament en la conservació d’A. tridens.
Dades publicades a GBIF (Doi: 10.15470/0u03uz)
Paraules clau: Chiroptera, Detecció acústica, Activitat nocturna, Masses d’aigua, Nord-oest d’Àfrica
Introduction
With more than 1,421 species (Simmons and Cirranellon, 2020), Chiroptera is the second most speciose mammalian order in the world, constituting about a quarter of all mammal species (Hutson et al., 2001; Wilson and Reeder, 2005). Bats have colonized all continents except Antarctica, with different levels of species richness (Owen, 1990; Georgiakakis et al., 2010; Curran et al., 2012; Fenton and Simmons, 2014; Weber and Cáceres, 2018). Bat species richness peaks in equatorial zones and is lowest in desert areas (Ramos Pereira and Palmeirim, 2013; Monadjem et al., 2018). As for other North-western African countries (Ahmim and Oubaziz, 2017 for Algeria; Aulagnier et al., 2017 for Morocco), few data have been collected in Tunisia from sub-Saharan and Saharan areas (Dalhoumi et al., 2011; Puechmaille et al., 2012). The first data on southern Tunisian bats were published by Baker et al. (1974) and Cockrum (in Dalhoumi et al., 2011). These authors recorded 6 bat species: Rhinolophus ferrumequinum, Asellia tridens, Eptesicus isabellinus, Pipistrellus kuhlii, Plecotus gaisleri and Myotis punicus. Later, Rebelo and Brito (2006) studied bat activity in several Tunisian desert localities but they did not provide details about species diversity. More recently, Bendjeddou et al. (2016) described a new species, Nyctinomus aegyptiacus, for the Tunisian fauna during a touristic visit. To date, only 47 records corresponding to 13 species of bats have been made for this area. In addition, little ecological information is available for the most common areas that cover around a third of the country. The lack of knowledge of bat fauna in Tunisia requires intensive surveys focusing mainly on habitat use in this harsh environment.
Bat acoustic surveys are standard in Europe and recommended for surveillance and monitoring of many species (Battersby, 2010). Acoustic monitoring in Northern Africa only started in the last decade (Dietz et al., 2007; Liéron et al., 2008; Benda et al., 2010; Dieuleveut et al., 2010; Disca et al., 2014), including surveys in Tunisia by Rebelo and Brito (2006) and Puechmaille et al. (2012). More recently, Dalhoumi et al. (2014, 2016, 2019) studied bat activity and habitat use in Tunisian localities and showed that acoustic monitoring is useful to study bats in this type of ecosystems as it can more accurately describe the species richness than methods such as mist-netting and cave monitoring. Due to logistic constraints, acoustic monitoring was often limited to only recording bat activity during the first part of the night, however, modern automatic bat detectors are now widely available and can record bat calls overnight (e.g. Adams et al., 2012).
The goal of this study was to obtain information on bat species richness and habitat use in a desert area in Tunisian. We recorded bat activity in eight sites at Dghoumes National Park during the nursing season, when bats are most active. This survey aimed to assess activity patterns of bats at Dghoumes National Park and identify their most important habitats. Understanding bat activity and diversity could provide information relevant to the conservation of several species within and outside the limits of the park.
Material and methods
Study area
Dghoumes National Park (34° 01′ N–34° 06′ N, 8°28′ E–8°39′ E) is located 30 km east of Degueche (Tozeur governorate, southwest Tunisia) (fig. 1). The park covers approximately f 8,000 ha. It borders with Chott el-Jérid in the south and the mountain ridge of Jebel Morra, Jebel El Kebiriti and Jebel Taferma in the north. The southern flank of the mountain range is cut through by wadis (Tamaxid, El Akreb, El Behim, El Kebiriti, Bou Moussa, El Oussif) that flow to Chott el-Jérid across a broad sandy plain (3,800 ha). The park has two small hillside lakes (Marzoug and Tamxid basins) and several permanent ponds (Sassia pond, El Kebiriti pond, and others). The bioclimate is arid with cool winters. The annual rainfall is about 100 mm/year. The average temperature ranges from -3 °C in winter to more than 50 °C in the summer. Evaporation reaches 1,200 mm/year. The sirocco wind blows for more than 50 days/year, especially in the summer (Abdelkader Chetoui, pers. comm.). The dominant biome is steppe with herbaceous vegetation such as Suaeda mollis, Zygophyllum album, Salsola tetrandra, Traganum nudatum, Artemisia campestris, Rhus tripartitum, Periploca laevigata and Retama retam, and sparse trees of Acacia tortilis raddiana and Tamarix aphylla. The main terrestrial mammals are Ctenodactylus gundi, Hyaena hyaena, Lepus capensis, Ammotragus lervia and reintroduced Oryx dammah and Gazella dorcas (Jebali, 2012).

Fig. 1. Study sites of a bat community in Dghoumes National Park (southwest Tunisia): 1, main park entrance; 2, Tamxid basin; 3, Marzoug basin; 4, El Kebiriti pond; 5, Sassia pond; 6, Jmal Ribeh pond; 7, El Oussif wadi; 8, plain. Fig. 1. Emplazamientos de estudio de la comunidad de murciélagos en el Parque Nacional de Dghoumes (suroeste de Túnez): 1, entrada principal del parque; 2, cuenca de Tamxid; 3, cuenca de Marzoug; 4, estanque El Kebiriti; 5, estanque Sassia; 6, estanque Jmal Ribeh; 7, Uadi El Oussif; 8, llanura.
Data collection and species identification
From June 16 to 24, 2018, we studied bat activity at eight sites of Dghoumes National Park (fig. 1): 1) at the main entrance to the park, which is illuminated by solar white street lamps (34° 2′ 30.62″N, 8° 33′ 51.95″E); 2) at Tamxid basin (34° 3′ 53.93″ N, 8° 31′ 13.27″ E) with a wet sludge; 3) at Marzoug basin (34° 3′ 57.73″ N, 8° 29′ 38.31″ E) with a dry sludge; 4) at El Kebiriti pond (34° 3′ 57.19″ N, 8°33′ 8.42″ E); 5) at Sassia pond (34° 4′ 15.44″ N, 8° 31′ 13.24″ E); 6) at Jmal Ribeh pond (34° 4′ 14.80″ N, 8° 29′ 55.83″ E) surrounded by phragmites and sunk into high banks; 7) at the dry wadi El Oussif (34° 2′ 53.24″ N, 8° 33′ 31.56″ E) with some retama bushes; and 8) at a plain (34° 2′ 55.13″ N, 8° 33′ 59.12″ E) characterized by sparse vegetation and five small water bodies.
Each site was surveyed for one night under similar hot and dry weather conditions. We used automatic echolocation call recordings at fixed points starting 30 minutes before sunset and lasting 30 minutes after sunrise. A Song Meter SM2BAT+ recorder (Wildlife Acoustic, Inc. Concord, Massachusetts, U.S.A.) was connected to a BMX-US1 ultrasonic microphone by a 3 m cable (Biotope, France). Real-time recordings were made at a sampling frequency of 384 kHz and an activation frequency of -18bB and 8 Hz. Recorded sequences were unzipped, analyzed and split into 5-second sequences with Kaleidoscope (Wildlife Acoustic, Inc. Version 4.1.0a). Only 5-second sequences containing bat calls were analyzed using the real-time analysis software BatSound, v.3.10 (Petterson Elektronik AB) for spectrogram analyses. We used a simple frequency of 38,400 samples/s, 16 bits/sample and selected 512 pt FFT with a Hamming window for analysis (Russ, 1999). Recorded calls, including feeding and drinking buzzes, were manually identified to species level using shape and other call parameters (Barataud, 2012). Echolocation calls of Tadarida teniotis, Eptesicus isabellinus, Pipistrellus kuhlii and Rhinopoma cystops were identified using reference calls collected in various sites in Tunisia (Dalhoumi et al. (2014, 2016, 2019). Asellia tridens uses a typical signal with constant frequency at 108-122 kHz (Jones et al., 1993). Vansonia rueppellii was identified by shallow FM/QCF calls at a frequency of maximum energy of 53.6 ± 1.3 kHz (Disca et al., 2014). A specimen of this species has been previously mist-netted at the entrance of the park and recorded after release during a visit on April 2, 2010. Additionally, temperature and hygrometry were recorded every 30 minutes during the nights of June 17-18 and 21-22 using a C.A. 846 pocket thermo-hygrometer (Chauvin Arnoux, Taiwan; precision ± 0.5 °C and ± 2.5 % RH) for potential bat roosts in caves, crevices and buildings, and visually counted emerging or resting individuals.
Statistical analyses
Bat activity was estimated by counting the number of passes (Russo and Jones, 2003). A bat pass was defined as a single or several calls emitted by a single animal during a 5 s interval (Barataud, 2012). Within each night, we considered 19 periods of 30 minutes to investigate activity patterns; the number of bat passes recorded during each period was used as a proxy of bat activity. Call recordings were used to calculate species richness at each site. Bat community analysis was performed using the number of bat passes. We used Kruskal-Wallis tests, followed by post-hoc multiple comparison tests, and Friedman tests to assess differences in bat activity among sites and periods. Statistical analyses were performed with Tanagra v.5, ’97 edition (Rakotomalala, 2005).
Results
We recorded 50,394 passes of six species at six of the eight sites surveyed in Dghoumes National Park (table 1, dataset published through GBIF (Doi: 10.15470/0u03uz)); no bat calls were recorded at the dry El Oussif Wadi (site 7) or on the plain (site 8). Even when sites without bat activity were excluded, this acoustic survey revealed a significant difference in activity between sites (Kruskal–Wallis test: H = 17.74; df = 5; P = 0.003). Most calls were recorded near the street lamps (site 1, 25.7 % of total bat passes), followed by activity at the Tamxid basin (site 2, 22.0 %), El Kebiriti pond (site 4, 20.7 %), Sassia pond (site 5, 14.4 %), Marzoug basin (site 3, 10.7 %), and Jmal Ribeh pond (site 6, 6.5 %). Differences were only significant between streetlamps, Tamxid basin, El Kebiriti pond, Sassia pond and Jmal Ribeh pond (p < 0.001 for the four paired comparisons; the post-hoc multiple comparison tests are very conservative).

Table 1. Number of bat passes, species richness and indexes recorded at eight sites of Dghoumes National Park (southwest Tunisia) in June 2018 (one night per site). Tabla 1. Número de pasos de murciélagos, riqueza de especies e índices registrados en ocho emplazamientos del Parque Nacional de Dghoumes (suroeste de Túnez) en junio de 2018 (una noche por emplazamiento).
Three species were recorded near the street lamps at the park entrance (site 1) with Pipistrellus kuhlii more commonly recorded (96.4 % of passes) than Eptesicus isabellinus and Asellia tridens. Four species were recorded at each water body (sites 2, 6): E. isabellinus, P. kuhlii, A. tridens and Tadarida teniotis, the latter being the least recorded. E. isabellinus, which was the only species that was mist-netted, and P. kuhlii were by far the most active. A. tridens was mainly recorded at temporary water bodies. Rhinopoma cystops was recorded at three water bodies (sites 2, 4 and 5), and Vansonia rueppelli was very rarely detected at two water bodies (sites 3, 6) (table 1).
Bats were most active overnight when temperature declined, and hygrometry increased (fig. 2) with a significant difference in activity between periods (Friedman test: H = 15.21, df = 5, P = 0.009). P. kuhlii (29,005 passes) and E. isabellinus (19,164 passes) were active overnight at water bodies and emitted feeding and drinking buzzes; differences between periods were significant (H = 86.15, df = 7, P < 0.001; H = 106.46, df = 7, P < 0.000, respectively). P. kuhlii displayed three periods of high activity: just after sunset near water bodies; three to four hours after sunset, mainly near the street lamps; and just before sunrise near the street lamps and water bodies (fig. 3A). E. isabellinus exhibited a similar pattern except that its activity was low just after sunset, and was very low overnight near the street lamps (fig. 3B). A. tridens was active late after sunset (fig. 4), activity peaked 1-2 hours after sunset and decreased slightly until sunrise. R. cystops was active soon after sunset (5 to 20 min) and was irregularly active until three hours before sunrise (fig. 4). The numbers of passes of the two remaining species were too low to determine a pattern.
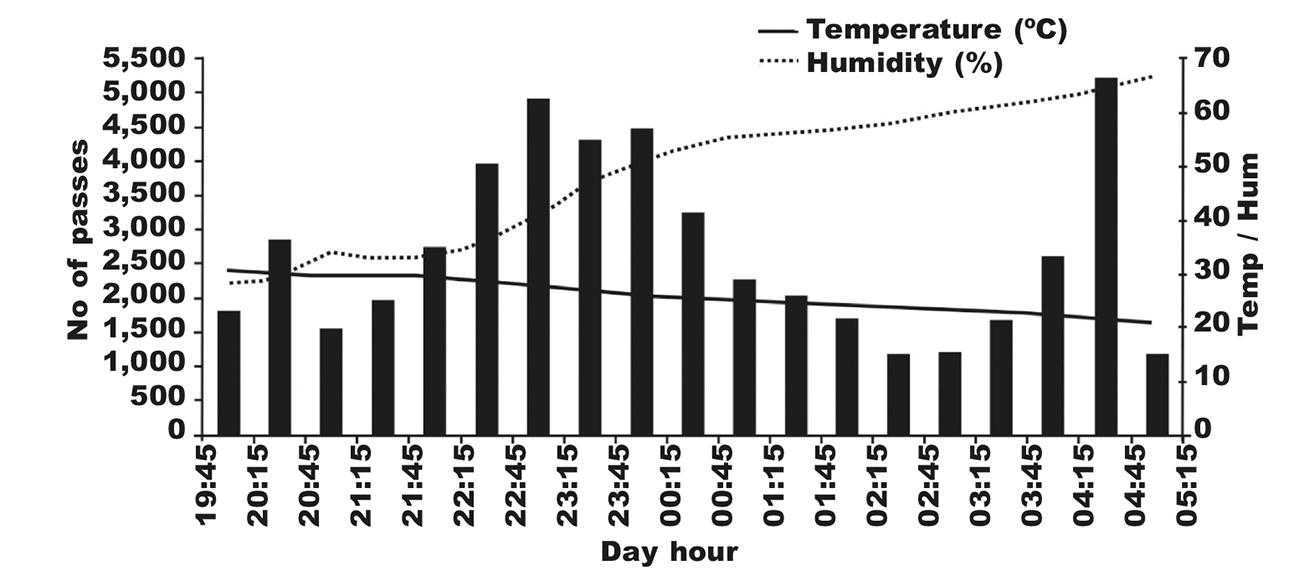
Fig. 2. Overnight activity pattern of bats for each 30–minute period starting after sunset in Dghoumes National Park (southwest Tunisia): sum of number of passes recorded at six sites (one night per site) from June 16 to 24, 2018. Fig. 2. Patrón de actividad nocturna de los murciélagos en periodos de 30 minutos a partir de la puesta del sol en el Parque Nacional de Dghoumes (suroeste de Túnez): suma del número de pasos registrados en seis emplazamientos (una noche por emplazamiento), del 16 al 24 de junio de 2018.

Fig. 3. Overnight activity pattern of Pipistrellus kuhlii (A) and Eptesicus isabellinus (B) for each 30–minute period starting after sunset at six sites of Dghoumes National Park (southwest Tunisia) in June 2018 (one night per site). Fig. 3. Patrón de actividad nocturna de Pipistrellus kuhlii y Eptesicus isabellinus (B) en periodos de 30 minutos a partir de la puesta del sol en seis emplezamientos del Parque Nacional de Dghoumes (suroeste de Túnez) en junio de 2018 (una noche por emplazamiento).

Fig. 4. Overnight activity pattern of Rhinopoma cystops, Asellia tridens, Tadarida teniotis and Vansonia rueppellii for each 30–minute period starting after sunset in Dghoumes National Park (southwest Tunisia) in June 2018 (one night per site). Fig. 4. Patrón de actividad nocturna de Rhinopoma cystops, Asellia tridens, Tadarida teniotis y Vansonia rueppellii en periodos de 30 minutos a partir de la puesta del sol en el Parque Nacional de Dghoumes (suroeste de Túnez) en junio de 2018 (una noche por emplazamiento).
Our visual bat census recorded 111 individuals of R. cystops emerging from the Jebel Morra cave on June 19, 2018, and 54 individuals of A. tridens resting in the dome of the Ecomuseum exhibition room on June 23, 2018.
Discussion
Bat richness was lower than that in similar studies in central and northern Tunisia (11 species in Jebel Mghilla National Park and Mastouta-Bishshouk region, Dalhoumi et al., 2014, 2019). Our results reflect the difficulty in detecting bat fauna in deserts even in areas where water is available throughout the year.
The bat assemblage recorded in this study included three Mediterranean (P. kuhlii, E. isabellinus and T. teniotis) and three Saharan bat species (R. cystops, A. tridens and V. rueppellii). Previously reported in this park (Puechmaille et al., 2012), P. kuhlii is widespread in Tunisia (Dalhoumi et al., 2011) and in northern Sahara (Happold and Happold, 2013). The ranges of E. isabellinus and T. teniotis only reach the Saharan border (Happold and Happold, 2013), and are possibly restricted to Chott el-Jérid in southwestern Tunisia. R. cystops and A. tridens, both widely distributed in the Sahara (Happold and Happold, 2013), were also reported in this Park by Gharaibeh (1997) and Puechmaille et al. (2012). V. rueppellii is considered rare in the Sahara (Happold and Happold, 2013) and only one specimen was collected by G. Nobis near Matmata, Tunisia (east of Chott el-Jérid) (Dalhoumi et al., 2011).
The bat assemblage in Dghoumes National Park could perhaps be expanded by one species, Myotis punicus, as it was previously reported close by in Nefta (Wandeler, 1967). Future studies in the area should aim at finding R. ferrumequinum, Otonycteris hemprichii and Plecotus gaisleri, which were reported in arid areas to the north and south-east of the Chott region in Tunisia (Dalhoumi et al., 2011; Puechmaille et al., 2012), and from similar zones of neighbouring countries (Kowalski and Rzebik-Kowalska, 1991; Ahmim and Oubaziz, 2017; Aulagnier et al. 2017). However, these species are difficult to detect during a short survey with a Song Meter detector (SM2+) because they emit signals with low intensity (Holderied et al., 2011; Barataud, 2012); this detector has a significant rate of attenuation with distance (Adams et al., 2012). Nyctinomus aegyptiacus was only reported in Ksar Ghilane (south Tunisia), where a dead specimen was identified (Bendjeddou et al., 2016); the echolocation calls of this bat have not been described previously in North Africa. Future studies should also aim at building high-quality echolocation call libraries.
In this survey, we only recorded bat echolocation calls at water bodies and street lamps. Our data confirm the strong affinity that bats have to water bodies in desert areas (Williams and Dickman, 2004; Rabe and Rosenstock, 2005; Rebelo and Brito, 2006; Razgour et al., 2010; Adams and Kwiecinski, 2018). Bats use these places to forage and drink, as in other Mediterranean areas (Russo and Jones, 2003; Di Salvo et al., 2009; Lisón and Calvo, 2011, 2014; Dalhoumi et al., 2014, 2016). The main entrance to the park is the only lit area in 20 km; the white light attracts some insects (Rowse et al., 2016) that may be preyed by bats such as Pipistrellus spp. (Haffner and Stutz, 1985-86; Blake et al., 1994; Lacoeuilhe et al., 2014). This type of lighting could deter other local fauna that also depend on the water bodies in the area. Following Voigt et al. (2018), who considered that all external artificial light should be avoided at night, we suggest that the present lamps should be replaced by lamps with a colour temperature < 2,700 K. This would ensure that the area is sufficiently lit for human visitors while simultaneously decreasing disturbance to the local fauna.
In Dghoumes National Park, the activity of most species of bats, mainly recorded as feeding and drinking buzzes, started at dusk near water bodies. Similarly, Griffiths (2013) and Russo et al. (2016) reported drinking buzzes only early on in the night, suggesting that bats cover their water requirements as soon as they emerge from their roost. For P. kuhlii, this first peak of activity was similar at water bodies in Bou Hedma National Park (central Tunisia, Dalhoumi et al., 2017) and in Algerian capture records (Gaisler and Kowalski, 1986). In our study, activity peaked for the second time 3h 30′ after sunset and for the third time one hour before sunrise. In the Negev desert, Razgour et al. (2011) also recorded three peaks of activity for P. kuhlii, with the two first peaks occurring slightly later in the night.
Foraging activity of E. isabellinus was concentrated in around several water bodies (Tamaxid basin, Sassia pond and El Kebriti pond) until midnight. It decreased shortly afterwards and ended at sunrise. This pattern, including a maximum soon after sunset, is in accordance with previous records of early night foraging (Brosset, 1955; Gaisler and Kowalski, 1986; Pérez Jordá, 1995). Contrary to our previous findings in Bou Hedma National Park (Dalhoumi et al., 2017), no significant activity was recorded near street lamps, suggesting they are not a swarming place (Altringham, 2011).
Finally, the activity of A. tridens peaked late after sunset and stopped before sunrise. This pattern should be confirmed as this species is known to be tolerant to light when roosting; it has been sighted flying in broad daylight in Morocco (Aulagnier et al., 2017), and reported to emerge before dusk in Algeria (Kowalski and Rzebik-Kowalska, 1991).
Conservations perspectives
The activity pattern of the six bat species recorded in Dghoumes National Park confirmed the importance of permanent water bodies for Mediterranean or desert bats. Considering the fast changing environmental conditions that we are facing, the rapid expansion of the desert, and the importance of environmental education that attracts visitors to this area, we suggest creating artificial ponds along the main wadi of the park and changing the type of lighting at the entrance to the park. These changes would encourage local bat fauna, and potentially other species, to use Dghoumes National Park as a refuge. Additionally, as the Ecomuseum hosts the most important colony of A. tridens known to date in Tunisia, and is jeopardized when the exhibition room is open to the public, we suggest restricting access to areas where the colony is roosting by fitting a false ceiling. Our recommendations are in line with the expectations of the World Bank donation to implement a series of conservation actions that better protect the fauna at Dghoumes National Park.
Acknowledgements
Bats were sampled and collected under Permit No. 1257 issued by the Directorate-General for Forests, Tunis. Chokri Issaoui manager of Dghoumes National Park and forest rangers provided great support and help in the field. Abdelkader Chitoui shared unpublished data, Hibat-Ellah Loumassine counselled on the identification of some bat calls, and anonymous reviewers helped to improve this manuscript by their comments. We are grateful to Eurobats for supplying us with the bat detectors.
















